Ultrasonic vocalizations as a tool for research on emotion and motivation in rodents
Markus Wöhr & Rainer K.W. Schwarting
Behavioural Neuroscience, Experimental and Physiological Psychology, Philipps-University of Marburg
Gutenbergstr. 18, 35032 Marburg, Germany
Mice and rats are the most common laboratory animals utilised in biomedical research, neuroscience and experimental psychology. These species have extensively been used to study basic mechanisms underlying emotional and motivational functions. Such research usually relies on overt behavioural measures, like approaching (or working for) an appetitive stimulus, or avoiding (or fleeing from) an aversive one. Besides, the presumed motivational and affective state is usually inferred from physiological measures, like heart rate, corticosterone excretion, or brain activity. In addition, or even extension to these measures, substantial information about the animal under study can also be obtained by gauging its vocalization, most of which often occurs in the ultrasonic range. Such ultrasonic vocalizations (USV) are behaviourally important for at least two aspects: For one, they can serve as a measure of the animal’s current affective state, and secondly, they seem to be an important communicative variable, which has to be taken into account when analysing social aspects of such species (mating, nursing, play, aggression, defence etc.).
Data on USV in rodents were reported as early as 1954. Anderson (Anderson, 1954) observed that adult laboratory rats emit calls at frequencies around 23-28 kHz when socially isolated. Two years later, Zippelius and Schleidt published their important discovery that infant mice produce USV when separated from their mother and litter (Zippelius et al., 1956). Since then, several studies were performed showing that measuring USV can provide new insights into emotion and motivation as well as communication in mice and rats.
Mouse
Three USV classes are currently known in mice. Their occurrence is dependent on the animal’s age and sex. Isolation-induced USV are emitted by mouse pups during the first two weeks of life when separated from mother and littermates. Interaction-induced USV occur mostly during social investigation in juvenile mice aged three to four weeks, at least in males. In females, however, they also occur in older individuals. Finally, female-induced USV are uttered by adult male mice when exposed to females or female urine (for review see: Portfors & Perkel, 2014; Wöhr & Schwarting, 2013).
Isolation-induced ultrasonic vocalizations
Affective state: Following Konrad Lorenz Zippelius and Schleidt (1956) named isolation-induced USV “Pfeifen des Verlassenseins” (“whistles of loneliness”), implying a negative affective state. Despite the fact that a wealth of evidence in support of the affective hypothesis has been accumulated since then, there is an ongoing scientific dispute on whether such isolation-induced USV reflect a negative affective state or whether they are simply the byproduct of a thermoregulation process (Blumberg & Sokoloff, 2001). Strongest argument in favour of the affective hypothesis is the fact that anxiolytic compounds such as benzodiazepines and other positive modulators of GABA-receptors reduce the production of isolation-induced USV (Benton & Nastiti, 1988; Fish et al., 2000; Nastiti et al., 1991; Takahashi et al., 2009). Also, it has been shown that mice bred for high anxiety-related behaviour on the elevated plus maze emit more isolation-induced USV than less anxious mice (Krömer et al., 2005; Kessler et al., 2011).
Communicative function: Zippelius and Schleidt (1956) suggested that isolation-induced USV serve communicative purposes, since they have observed that mothers leave the nest to retrieve vocalizing pups scattered outside the nest, whereas no retrieval behaviour was seen in response to pups that have been anesthetized and hence did not emit isolation-induced USV. Later, it shown by means of playback experiments that indeed isolation-induced ultrasonic vocalizations elicit maternal search and retrieval behaviour (Ehret, 1987; Ehret & Haack, 1981; Sewell, 1970; Smith, 1976; Wöhr et al., 2008a).
In line with their communicative function, isolation-induced USV are increasingly used as a measure for the quantification of communication deficits in mouse models of neuropsychiatric disorders characterized by social and communication deficits (Scattoni et al., 2009; Silverman et al., 2010). There are a number of genetic mouse models of autism, speech impairment and related disorders displaying a reduction of isolation-induced USV (Chadman et al., 2008; Enard et al., 2009; Fujita et al., 2008; Gaub et al., 2010; Groszer et al., 2008; Jiang et al., 2010; Kurz et al., 2010; Laviola et al., 2006; Moles et al., 2004; Nakatani et al., 2009; Shu et al., 2005; Winslow & Insel, 2002; Young et al., 2010), while other mouse models, e.g. the BTBR T+tf/J inbred strain mouse model of autism, are characterized by an unusual repertoire of isolation-induced USV subtypes (Scattoni et al., 2008) and a limited ability and/or reduced motivation to adjust their emission to different social contexts (Wöhr, 2015). Detailed spectrographic analyses have further shown that some mouse models of autism and related disorders are further characterized by altered call clustering and reduced temporal organization of USV emission (Ey et al., 2013; Mosienko et al., in press; Schmeisser et al., 2012; Wöhr, 2014).
Interaction-induced ultrasonic vocalizations
Affective state: Recently, Panksepp et al. (2007) reported that also juvenile mice produce USV. They found high rates of USV during social interactions of four week old mice. As the production of interaction-induced USV occurred during the play period and no aggressive behaviour was observed, it is tempting to speculate that they reflect a positive affective state alike rat 50-kHz USV that also occur during play behaviour (Knutson et al., 1998; Lukas & Wöhr, 2015). It is remarkable that emission of interaction-induced USV declines rapidly with sexual maturity – at the time when male mice start to display aggressive behaviour. Then, male mice begin to emit female-induced USV. In females, however, interaction-induced USV persist into adulthood (Moles et al., 2007) and were found to be a particularly sensitive measure for autism-related communication deficits, e.g. in the Shank2 mouse model of autism (Ey et al., 2013; Schmeisser et al., 2012).
Communicative function: It was suggested that interaction-induced USV help to maintain social contact, since their occurrence is positively associated with social investigation behaviour (Panksepp et al., 2007). In line with an affiliative function, Scattoni et al. (2013) reported that the lack of social investigation behaviour in the BTBR T+tf/J autism model is paralleled by a lack of interaction-induced USV in juveniles. Similar findings were obtained in adult mice (Scattoni et al., 2011).
Female-induced ultrasonic vocalizations
Affective state: In adult mice, high rates of USV were found in males when courting and copulating with females (Sewell, 1967). As shown by Whitney et al. (1974), female urine alone, i.e. in the absence of a female, is sufficient for eliciting male USV. No vocalization response, however, was detected when male mice were exposed to male mouse urine or female urine from rats or humans (Wang et al., 2008; Whitney et al. 1974). The production of male USV in response to females has song-like characteristics, including different syllable types and a temporal sequencing that includes the emission of repeated phrases (Holy & Guo, 2005). Interestingly, female-induced USV are highly sensitive to important social factors such as social status (D’Amato, 1991; Nyby et al., 1976) and previous heterosexual contact (Dizinno et al., 1978; Maggio et al., 1983; Nyby et al., 1977; Nyby et al.1983; Roullet et al., 2011; Sipos et al., 1992; Sipos et al.,1995). The notion that female-induced USV reflect a positive affective state alike rat 50-kHz USV (Panksepp & Burgdorf, 2003) is supported by the fact that USV in adult male mice can also be induced by amphetamine (Wang et al., 2008).Consistently, it was found that male mice displaying a mania-like phenotype emit particularly high rates of female-induced USV (Malkesman et al., 2010).
Communicative function: As shown in devocalization and playback studies, female-induced USV serve an important communicative function, namely to attract females similar to bird song (Hammerschmidt et al., 2009; Pomerantz et al., 1983). Unlike birds, however, mice do not require auditory input and do not learn female-induced USV through imitation. In fact, deaf mice were found to emit female-induced USV that did not differ from the ones emitted by controls able to hear (Hammerschmidt et al., 2012; Mahrt et al., 2013). As for isolation- and interaction-induced USV, reduced levels of female-induced USV were reported in mouse models of autism (Jaiman et al., 2008; Radyushkin et al., 2009; Wöhr et al., 2011b).
Rat
Dependent on sound frequency, call lengths and frequency modulation, at least three different classes of USV can be identified in the rat. Their occurrence differs dependent on animal age, and physical, or psychological demands of the environment (for review see: Brudzynski, 2013; Wöhr & Schwarting, 2013).
Isolation-induced ultrasonic vocalizations
Affective state: Rat pups typically exhibit USV around the 40-kHz level in response to several distressing situations, like separation from their litter and mother (Hofer, 1996; Hofer et al., 1978), or when ambient temperature drops (Blumberg et al., 1996). Considering the affective value of such experiences it was postulated that infant calling reflects a negative affective state. Indeed, isolation-induced USV appear to be a valid index of pup anxiety, since the rate of calling can be attenuated by anxiolytic drugs (Hofer, 1996; Insel et al., 1986; Kehne et al., 2000; Olivier et al., 1998a; Olivier et al., 1998b; Vivian et al., 1997) and, as in mice, rats bred for high anxiety-related behaviour on the elevated plus maze emit more isolation-induced USV than less anxious rats (Wigger et al., 2001). Furthermore, distress calls also act as a main elicitor of anogenital licking (Brouette-Lahlou et al., 1992). Such licking serves a vital function, since non-licked pups cannot defecate, and do not survive. Interestingly, maternal care can affect the development of emotionality. Adult rats, which had been licked more often during infancy, show less anxiety-related behaviour in response to aversive stimuli than less frequently licked rats (Caldji et al., 1998). In accordance with this finding, it was shown that frequently licked animals emitted less isolation-induced USV, i.e. less anxiety-related behaviour, in infancy than rarely licked animals (Wöhr & Schwarting, 2008a).
In contrast to simple USV measures like occurrence and frequency range, much less is known about the spectrographic structure of such calls. Until recently, most research on USV in rats has been conducted with simple bat detectors set on limited frequency ranges. Since the emission of these 40-kHz calls is characterized by a high variability, calls occurring at different frequency ranges might be missed. Therefore, researchers have begun to look at spectrographic characteristics in more detail. Brudzynski et al. (1999) were able to classify infant calls according to ten categories by using a recording system that is capable of monitoring a wide range of frequencies. Spectrographic analysis revealed a developmental trend emphasizing those call characteristics, which are important for pup survival (Brudzynski et al., 1999). More recently, detailed spectrographic analysis provides evidence that pharmacological treatment can selectively affect certain call patterns rather than USV production in general (Barron et al., 2005). Furthermore, it was shown that call characteristics, like amplitude, frequency modulation and bout structure, are related to the amount of maternal care experienced during early life (Wöhr & Schwarting, 2008a).
Communicative function: Isolation-induced USV seem to have an important value for pup survival. As isolation-induced USV in mice, rat 40-kHz USV can elicit maternal search and retrieval behaviour as demonstrated by means of playback studies (Allin et al., 1972; Smotherman et al., 1974; Wöhr & Schwarting, 2008a).
Fear-induced ultrasonic vocalizations
Affective state: Adult rats produce two different types of USV. These calls have been classified primarily on the basis of their sound frequency as low and high frequency USV. Low frequency vocalizations, so called 22-kHz USV, are within a frequency range of 18 – 32 kHz. The calls have a sound pressure level of 65–85 dB and call durations of approximately 300–4000 ms (Borta et al., 2006; Brudzynski, 2001; Sales et al., 1974; van der Poel et al., 1989; Wöhr et al., 2005). High frequency vocalizations, so called 50-kHz USV, are within a frequency range of 32–96 kHz, with short durations of 30–50 ms (Brudzynski & Pniak, 2002; Kaltwasser, 1990; Sales et al., 1974; White et al., 1990). Increasing evidence from ethological, pharmacological, and brain stimulation studies suggest that these USV are critically dependent on situational factors and experience, and may represent distinct affective states of the subject. Low-frequency 22-kHz USV are emitted by adult rats when exposed to predators (Blanchard et al., 1991), or other aversive stimuli, like startling noises (Kaltwasser, 1991), or unescapable foot-shocks (Antoniadis et al., 1999; Borta et al., 2006; Vivian et al., 1993; Wöhr et al., 2005). Furthermore, 22-kHz USV are emitted during intermale aggression (Vivian et al., 1993).
Such 22-kHz USV are not only emitted during the actual aversive event, but also in response to stimuli associated with such experiences (Antoniadis et al., 1999; Borta et al., 2006; Cuomo et al., 1988; De Vry et al., 1993; Molewijk et al., 1995; van der Poel et al., 1989; Wöhr et al., 2005). It is thus assumed that these calls reflect a negative affective state akin to anxiety and depression. Consistently, it was shown that aversive experiences during early development lead to increased emission of 22-kHz USV in adulthood (Yee et al., 2012a). In addition, it was found that measuring 22-kHz USV helps to reveal effects of early life stress that are not detectable by means of classical behavioural measures, such as freezing behaviour (Yee et al., 2012b).
However, little information is available upon which acoustic parameter is carrying the strength of the signal. Interestingly, it was shown that not only the number of calls emitted is related to the intensity of an emotional state in the rat, but that latencies to utter vocalizations, call lengths and loudness can also reflect the intensity of an aversive emotional state (Wöhr et al., 2005). Furthermore, rats, which were defined as highly anxious based on the time spent in the open arms of the elevated plus maze, tended to vocalize more often than rats displaying low anxiety-like behaviour, and exhibited a higher peak frequency (Borta et al., 2006).
Communicative function: Blanchard et al. (1991) suggested that 22-kHz USV serve as alarm calls to warn conspecifics about danger. They observed that 22-kHz USV production in response to a predator, a cat, is dependent on the presence of conspecifics. In absence of such an audience, no 22-kHz USV were observed when a rat was exposed to a cat. Although an audience effect was not replicated in standardized laboratory conditions (Wöhr & Schwarting, 2008b), there is evidence that 22-kHz USV can induce anxiety-related behaviour such as freezing in the recipient, supporting the notion that 22-kHz USV indeed serve an alarm function. By means of playback experiments it was shown that 22-kHz USV induce locomotor inhibition (Wöhr & Schwarting, 2007) and neuronal activity in brain areas implicated in fear regulation, such as amygdala and central grey (Parsana et al., 2012b; Sadananda et al., 2008). Recent findings indicate that rats are predisposed to form memory associations between 22-kHz USV and aversive stimuli (Bang et al., 2008; Endres et al., 2007; Furtak et al., 2007; Wöhr & Schwarting, 2010), possibly through auto conditioning (Kim et al., 2010; Parsana et al., 2012a), and that 22-kHz USV play an important role in the social transmission of fear (Kim et al., 2010). Finally, there is evidence that 22-kHz USV evoke an empathy-like state in the recipient – and by this means promote helping behaviour (Ben-Ami Bartal et al., 2011).
Interaction-induced ultrasonic vocalizations
Affective state: High-frequency 50-kHz USV occur in naturalistic
contexts including juvenile play (Knutson et al., 1998; Lukas &
Wöhr, 2015), tickling (Burgdorf & Panksepp, 2001; Mällo et al.,
2007; Panksepp & Burgdorf, 2000; Panksepp & Burgdorf, 2003;
Schwarting et al., 2007; Wöhr et al., 2009), social exploratory activity
(Brudzynski & Pniak., 2002), and mating behviour (McGinnis et al.,
2003; White et al., 1990). Since 50-kHz USV are also expressed during
anticipation of copulation (Bialy et al., 2000), play (Knutson et al.,
1998), food (Burgdorf et al., 2000), and electrical stimulation of the
medial forebrain bundle (Burgdorf et al., 2000), it has been postulated
that these calls are sensitive marker for unconditioned and conditioned
reward states (Knutson et al., 1999) and Panksepp and Burgdorf (2003)
assumed that these calls reflect a positive affective state akin to joy.
On the basis of these findings, 50-kHz USV are increasingly used as a
measure of positive affect in drug studies. It was repeatedly shown that
administration of amphetamine elicits high rates of 50 kHz USV in a
dose-dependent manner (Ahrens et al., 2009; Burgdorf et al., 2001;
Knutson et al., 1999; Natusch & Schwarting, 2010; Wintink et al.,
2001; Wright et al., 2010). Amphetamine-induced 50-kHz USV emission is
affected by the test context. Higher call rates are emitted when testing
is performed in test environments with fresh bedding material (Natusch
& Schwarting, 2010). Also, social context has an impact on
amphetamine-induced 50-kHz USV production (Wright et al., 2010). Wright
et al. (2010) performed a detailed spectrographic analysis of 50-kHz USV
elicited by amphetamine and identified 14 categories of 50-kHz USV.
They found that pair-tested rats produce a higher proportion of
frequency-modulated 50-kHz USV than individually-tested rats under both
amphetamine and saline conditions, and that amphetamine treatment alters
the call profile such that frequency-modulated 50-kHz USV became more
prominent while flat 50-kHz USV occurred less often. By means of cocaine
self-administration experiments, it was further demonstrated that
50-kHz USV can also serve as a useful marker for affective responses to
cocaine administration, anticipation and craving (Barker et al., 2010;
Maier et al., 2010). Finally, in line with the idea that 50-kHz USV
reflect a positive affective state and that amphetamine induces mania,
it was suggested that the amphetamine-induced increase in 50-kHz USV
could be used as readout for a mania-like state in rat models (Pereira
et al., 2014). In fact, Pereira et al. (2014) found that lithium, a mood
stabilizer typically used to treat bipolar disorder, blocks the
amphetamine-induced increase in 50-kHz USV.
As the production of aversive 22-kHz USV, emission of 50-kHz USV is
characterized by huge inter-individual differences (Mällo et al., 2007;
Schwarting et al., 2007; Wöhr et al., 2009). In tickling experiments, it
was found that not all rats emit 50-kHz USV – some rats do not emit USV
at all, others even emit 22-kHz USV (Mällo et al., 2007; Schwarting et
al., 2007; Wöhr et al., 2009). Such inter-individual differences are
considered predictive of stable individual traits in tests for social
behaviour, anxiety and depression. For instance, by means of an
innovative ambiguous cue interpretation paradigm for assessing
optimistic decisions in rats, Rygula et al. (2012) found that rats
emitting high rates of 50-kHz USV display an optimistic bias that was
not seen in rats emitting low 50-kHz USV rates. In subsequent studies,
it was then shown that rats displaying an optimistic bias are motivated
to gain reward (Rygula et al., 2015).
Moreover, rats can be selectively bred for low and high rates of 50-kHz
USV emission and such breeding is affecting social behaviour as well as
anxiety- and depression-related behaviour. Rats bred for low levels of
50-kHz USV spent less time in contact with conspecifics in adulthood
(Burgdorf et al., 2009) and did not show a preference for maternally
associated stimuli in infancy (Harmon et al., 2008), while rats bred for
high rates of 50-kHz USV displayed less anxiety-related behaviour as
indicated by more center entries in an open field in adulthood (Burgdorf
et al., 2009) and fewer isolation-induced USV in infancy (Harmon et
al., 2008). Consistently, rats bred for high anxiety-related behaviour
on the elevated plus maze were found to emit fewer 50-kHz USV during
juvenile play behaviour (Lukas & Wöhr, 2015).
Recently, inter-individual differences in the production of 50-kHz USV
were found to be linked with neurogenesis in the dentate gyrus of the
hippocampus (Wöhr et al., 2009; Yamamuro et al., 2010). While the number
of 50-kHz USV emitted during tickling was highly positively correlated
with hippocampal cell proliferation, a highly negative correlation
between 22-kHz USV and hippocampal cell proliferation was obtained.
Interestingly, neurogenesis in the hippocampus has been repeatedly
associated with affect regulation and psychopathology such as
depression. Thus, it is well known that aversive stimuli and events like
social defeat reduce hippocampal cell proliferation and that
hippocampal cell proliferation is necessary for the antidepressant
effects of selective serotonin reuptake inhibitors (Santarelli et al.,
2003). It is therefore remarkable that neurogenesis in the dentate gyrus
of the hippocampus is enhanced by tickling in rats that experienced the
tickling stimulation presumably as appetitive as indicated by high
numbers of 50-kHz USV (Wöhr et al., 2009; Yamamuro et al., 2010).
Hippocampal cell proliferation levels in rats that emitted high rates of
50-kHz USV during tickling were almost as double as high than in
non-tickled controls or in rats that emitted only some very few 50-kHz
USV during tickling (Wöhr et al., 2009).
Communicative function: Besides appetitive situations, 50-kHz USV
also occur after separation from conspecifics during short social
isolation. Rats taken out from their home cage and individually exposed
to a clean cage emit 50-kHz USV (Schwarting et al., 2007; Wöhr et al.,
2008b). Typically, 50-kHz USV emission is highest during the very first
minutes after separation from conspecifics (Wöhr et al., 2008b). For at
least two reasons it appears to be unlikely that the emission of 50-kHz
USV is novelty-induced. Firstly, 50-kHz USV emission is stable across
testing days, i.e. no habituation occurs (Wöhr et al., 2008b). Secondly,
not only the rat that has been taken out from the home cage starts to
emit 50-kHz USV, but also the rat that stays alone in the home cage
after the conspecific has been removed (Wöhr et al., 2008b). The fact
that separation from conspecifics elicits 50-kHz USV indicates an
affiliative communicative function of 50-kHz USV, namely to
(re)establish or to maintain social contact.
An affiliative communicative function of 50-kHz USV is also in line with
the observation that rats spent more time with conspecifics that
vocalize a lot than with those that display little emission of 50-kHz
USV (Panksepp et al., 2002). Furthermore, it was found that 50-kHz USV
promote and maintain rough and tumble play in juvenile rats (Himmler et
al., 2014). Consistently, deafening (Siviy & Panksepp, 1987) and
devocalizing (Kisko et al., 2015) rats reduces rough and tumble play.
Finally, Brudzynski and Pniak (2002) found that rats emit 50-kHz USV in a
dose-dependent manner when exposed to odor of conspecifics, indicating
that 50-kHz USV utterance is driven by potential social contact.
Subsequent playback studies further support the notion that 50-kHz USV
serve as social contact calls. While aversive 22-kHz USV induce
behavioural inhibition in the recipient, appetitive 50-kHz USV induce
social exploratory behaviour, both in male (Wöhr & Schwarting, 2007;
Wöhr & Schwarting, 2009) and female recipients (Willadsen et al.,
2014). For instance, in juvenile male rats, exposure of rats to 50-kHz
USV induces a three-fold increase in locomotor activity in comparison to
that induced in test phases without acoustic playback or phases where
an acoustic control stimulus was presented. The induced locomotor
activity was clearly directed towards the loudspeaker. During the 60 s
period of 50-kHz USV playback, juvenile male rats spent on average less
than 5 s away from the speaker, while more than 40 s near to it. Such a
preference was not observed during test phases without USV playback or
when an acoustic control stimulus was presented. Importantly, social
approach behaviour in response to playback of appetitive 50-kHz USV was
found to be dependent on active social interactions during the rough and
tumble play period and not seen in rats exposed to post-weaning social
isolation – yet again detectable after one additional week of social
housing (Seffer et al., in press).
The opposite behavioural responses elicited by playback of aversive
22-kHz USV and appetitive 50-kHz USV is paralleled by distinct patterns
of brain activation. While playback of aversive 22-kHz USV is followed
by an increase of neuronal activity in brain areas implicated in the
regulation of anxiety and fears, such as amygdala and central grey,
50-kHz USV inhibit neuronal activity in the amygdala (Parsana et al.,
2012b), while eliciting neuronal activity in the nucleus accumbens, a
key area for reward processing (Sadananda et al., 2008). Consistently,
it was recently shown that playback of appetitive 50-kHz USV elicits a
phasic release of dopamine in the nucleus accumbens and that the
magnitude of dopamine release is positively correlated with the strength
of social approach behaviour in the recipient (Willuhn et al., 2014).
Finally, a first pharmacological study shows that social approach
displayed in response to playback of 50-kHz USV can be used for studying
neurochemical mechanisms underlying social motivation and interest
(Wöhr et al., 2009). Endogenous opioids play an important role in the
regulation of social behaviours. In rats, particularly rough and tumble
play is affected by the administration of exogenous opioids. It was
consistently shown that the administration of low doses of
µ-opioid-receptor-agonists, such as morphine, increases rough and tumble
play, while a decrease in rough and tumble play was observed after
administration of µ-opioid-receptor-antagonists, such as naloxone
(Vanderschuren et al., 1997). In line with these findings it was found
that also social approach behaviour in response to playback of 50-kHz
USV is affected by the administration of exogenous opioids. In juvenile
as well as in adult rats, social approach behaviour was reduced after
naloxone treatment, but enhanced with morphine (Wöhr et al., 2009). In
addition to overt behavioural changes, ultrasonic calling in response to
playback of 50-kHz USV was affected by opioid ligands in juvenile rats.
While saline- and morphine-treated rats vocalized, no vocalization
response was detected in rats treated with naloxone (Wöhr et al., 2009).
This finding is in line with a study on ultrasonic communication in
µ-opioid-receptor-knockout mice. Typically, male mice display social
exploratory activity when exposed to interaction-induced USV emitted by
females (Wöhr et al., 2011a). Remarkably, adult male mice lacking the
µ-opioid-receptor, however, do not display social exploratory behaviour
under such conditions (Wöhr et al., 2011a). Together, this indicates
that an important feature of rodent social behaviour, namely ultrasonic
communication, is at least partially regulated by endogenous opioids.
Conclusion
Production of USV can be utilized to study the neuroanatomical and pharmacological basis of motivation and emotion. Behavioural responses to playback of USV can provide important insights into the social brain and help to elucidate genetic, neurochemical and neuroanatomical factors underlying neuropsychiatric disorders characterized by social deficits such as autism. The application of new, more sophisticated recording systems and sound analysis tools which enables the researcher to create detailed spectrograms provides a clear improvement for research on USV. By using these tools the investigator can detect critical differences between treatments or individual animals, which may not be detectable with standard behavioural measurements.
Further reading
For more detailed overviews on ultrasonic communication in rodents please see:
BRUDZYNSKI, S.M. (2013). Ethotransmission: communication of emotional states through ultrasonic vocalization in rats. Current Opinion in Neurobiology, 23, 310-317.
PORTFORS, C.V. & PERKEL, D.J. (2014). The role of ultrasonic vocalizations in mouse communication. Current Opinion in Neurobiology, 28, 115-120.
WÖHR, M. & SCHWARTING, R.K.W. (2013). Affective communication in rodents: ultrasonic vocalizations as a tool for research on emotion and motivation. Cell and Tissue Research, 354, 81-97.
There is also an overview in German available:
WÖHR, M. & SCHWARTING, R.K.W. (2010). Ultraschallkommunikation bei Nagern und ihre Bedeutung für Modelle neuropsychiatrischer Erkrankungen. Neuroforum, 4.10, 248-258.
References:
AHRENS, A.M., MA, S.T., MAIER, E.Y., DUVAUCHELLE, C.L. & SCHALLERT, T. (2009). Repeated intravenous amphetamine exposure: rapid and persistent sensitization of 50-kHz ultrasonic trill calls in rats. Behavioural Brain Research, 197, 205-209.
ALLIN, J.T. & BANKS, E.M. (1972). Functional aspects of ultrasound production by infant albino rats (Rattus norvegicus). Animal Behaviour, 20, 175-185.
ANDERSON, J.W. (1954). The production of ultrasonic sounds by laboratory rats and other mammals. Science, 119, 808-809.
ANTONIADIS, E.A. & MCDONALD, R.J. (1999). Discriminative fear conditioning to context expressed by multiple measures of fear in the rat. Behavioural Brain Research, 101, 1-13.
BANG, S.J., ALLEN, T.A., JONES, L.K., BOGUSZEWSKI, P. & BROWN, T.H. (2008). Asymmetrical stimulus generalization following differential fear conditioning. Neurobiology of Learning and Memory. 90, 200-216.
BARRON, S. & GILBERTSON, R. (2005). Neonatal Ethanol Exposure but not Neonatal Cocaine Selectively Reduces Specific Isolation-Induced Vocalization Waveforms in Rats. Behavior Genetics, 35, 93-102.
BARKER, D.J., ROOT, D.H., MA, S., JHA, S., MEGEHEE, L., PAWLAK, A.P. & WEST, M.O. (2010). Dose-dependent differences in short ultrasonic vocalizations emitted by rats during cocaine self-administration. Psychopharmacology, 211, 435-442.
BEN-AMI BARTAL, I., DECETY, J. & MASON, P. (2011). Empathy and pro-social behavior in rats. Science, 334, 1427-1430.
BENTON, D. & NASTITI, K. (1988). The influence of psychotropic drugs on the ultrasonic calling of mouse pups. Psychopharmacology, 95, 99-102.
BIALY, M., RYDZ, M. & KACZMAREK, L. (2000). Precontact 50-kHz vocalizations in male rats during acquisition of sexual experience. Behavioral Neuroscience, 114, 983-90.
BLANCHARD, R.J., BLANCHARD, D.C., AGULLANA, R. & WEISS, S.M. (1991). Twenty-two kHz alarm cries to presentation of a predator, by laboratory rats living in visible burrow systems. Physiology and Behavior, 50, 967-72.
BLUMBERG, M.S. & SOKOLOFF, G. (2001). Do infant rats cry? Psychological Review, 108, 83-95.
BLUMBERG, M.S. & STOLBA, M.A. (1996). Thermogenesis, myoclonic twitching, and ultrasonic vocalization in neonatal rats during moderate and extreme cold exposure. Behavioral Neuroscience, 110, 305-14.
BORTA, A., WÖHR, M. & SCHWARTING, R.K.W. (2005). Rat ultrasonic vocalization in aversively motivated situations and the role of individual differences in anxiety-related behavior. Behavioural Brain Research, 166, 271-80.
BROUETTE-LAHLOU, I., VERNET-MAURY, E. & VIGOUROUX, M. (1992). Role of pups’ ultrasonic calls in a particular maternal behavior in Wistar rat: pups’ anogenital licking. Behavioural Brain Research, 50, 147-54.
BRUDZYNSKI, S.M. (2001). Pharmacological and behavioral characteristics of 22 kHz alarm calls in rats. Neuroscience and Biobehavioral Reviews, 25, 611-617.
BRUDZYNSKI, S.M. (2013). Ethotransmission: communication of emotional states through ultrasonic vocalization in rats. Current Opinion in Neurobiology, 23, 310-317.
BRUDZYNSKI, S.M., KEHOE, P. & CALLAHAN, M. (1999). Sonographic structure of isolation-induced ultrasonic calls of rat pups. Developmental Psychobiology, 34, 195-204.
BRUDZYNSKI, S.M. & PNIAK, A. (2002). Social contacts and production of 50-kHz short ultrasonic calls in adult rats. Journal of Comparative Psychology, 116, 73-82.
BURGDORF, J., KNUTSON, B. & PANKSEPP, J. (2000). Anticipation of rewarding electrical brain stimulation evokes ultrasonic vocalization in rats. Behavioral Neuroscience, 114, 320-7.
BURGDORF, J., KNUTSON, B., PANKSEPP, J. & IKEMOTO, S. (2001a). Nucleus accumbens amphetamine microinjections unconditionally elicit 50-kHz ultrasonic vocalizations in rats. Behavioral Neuroscience, 115, 940-4.
BURGDORF, J. & PANKSEPP, J. (2001b). Tickling induces reward in adolescent rats. Physiology and Behavior, 72, 167-173.
BURGDORF, J., PANKSEPP, J., BRUDZYNSKI, S.M., BEINFELD, M.C., CROMWELL, H.C., KROES, R.A. & MOSKAL, J.R. (2009). The effects of selective breeding for differential rates of 50-kHz ultrasonic vocalizations on emotional behavior in rats. Developmental Psychobiology, 51, 34-46.
CALDJI, C., TANNENBAUM, B., SHARMA, S., FRANCIS, D., PLOTSKY, P.M., MEANEY, M.J. (1998). Maternal care during infancy regulates the development of fearfulness in the rat. Proceedings of the National Academy of Sciences of the United States of America, 95, 5335-5340.
CHADMAN, K.K., GONG, S., SCATTONI, M.L., BOLTUCK, S.E., GANDHY, S.U., HEINTZ, N. & CRAWLEY, J.N. (2008). Minimal aberrant behavioral phenotypes of neuroligin-3 R451C knockin mice. Autism Research, 1, 147-158.
CUOMO, V., CAGIANO, R., DE SALVIA, M.A., MASELLI, M.A., RENNA, G. & RACAGNI, G. (1988). Ultrasonic vocalization in response to unavoidable aversive stimuli in rats: effects of benzodiazepines. Life Science, 43, 485-491.
D’AMATO, F.R. (1991). Courtship ultrasonic vocalizations and social status in mice. Animal Behaviour, 41, 875-885.
DE VRY, J., BENZ, U., SCHREIBER, R. & TRABER, J. (1993). Shock-induced ultrasonic vocalization in young adult rats: a model for testing putative anti-anxiety drugs. European Journal of Pharmacology, 249, 331-339.
DIZINNO, G., WHITNEY, G. & NYBY, J. (1978). Ultrasonic vocalizations by male mice (Mus musculus) to female sex pheromone: experimental determinants. Behavioral Biology, 22, 104-113.
EHRET, G. & HAACK, B. (1981). Categorial perception of mouse pup ultrasound by lactating females. Naturwissenschaften, 68, 208.
EHRET G. (1987). Left hemisphere advantage in the mouse brain for recognizing ultrasonic communication calls. Nature, 325, 249-251.
ENARD, W., GEHRE, S., HAMMERSCHMIDT, K., HÖLTER, S.M., BLASS, T., SOMEL, M., BRÜCKNER, M.K., SCHREIWEIS, C., WINTER, C., SOHR, R., BECKER, L., WIEBE, V., NICKEL, B., GIGER, T., MÜLLER, U., GROSZER, M., ADLER, T., AGUILAR, A., BOLLE, I., CALZADA-WACK, J., DALKE, C., EHRHARDT, N., FAVOR, J., FUCHS, H., GAILUS-DURNER, V., HANS, W., HÖLZLWIMMER, G., JAVAHERI, A., KALAYDJIEV, S., KALLNIK, M., KLING, E., KUNDER, S., MOSSBRUGGER, I., NATON, B., RACZ, I., RATHKOLB, B., ROZMAN, J., SCHREWE, A., BUSCH, D.H., GRAW, J., IVANDIC, B., KLINGENSPOR, M., KLOPSTOCK, T., OLLERT, M., QUINTANILLA-MARTINEZ, L., SCHULZ, H., WOLF, E., WURST, W., ZIMMER, A., FISHER, S.E., MORGENSTERN, R., ARENDT, T., DE ANGELIS, M.H., FISCHER, J., SCHWARZ, J. & PÄÄBO, S. (2009). A humanized version of Foxp2 affects cortico-basal ganglia circuits in mice. Cell, 137, 961-971.
ENDRES, T., WIDMANN, K. & FENDT, M. (2007). Are rats predisposed to learn 22 kHz calls as danger-predicting signals? Behavioural Brain Research, 185, 69-75.
EY, E., TORQUET, N., LE SOURD, A.M., LEBLOND, C.S., BOECKERS, T.M., FAURE, P. & BOURGERON, T. (2013). The Autism ProSAP1/Shank2 mouse model displays quantitative and structural abnormalities in ultrasonic vocalisations. Behavioural Brain Research, 256, 677-689.
FISH, E.W., SEKINDA, M., FERRARI, P.F., DIRKS, A. & MICZEK, K.A. (2000). Distress vocalizations in maternally separated mouse pups: modulation via 5-HT(1A), 5-HT(1B) and GABA(A) receptors. Psychopharmacology, 149, 277-285.
FUJITA, E., TANABE, Y., SHIOTA, A., UEDA, M., SUWA, K., MOMOI, M.Y. & MOMOI, T. (2008). Ultrasonic vocalization impairment of Foxp2 (R552H) knockin mice related to speech-language disorder and abnormality of Purkinje cells. Proceedings of the National Academy of Sciences of the United States of America, 105, 3117-3122.
FURTAK, S.C., ALLEN, T.A. & BROWN, T.H. (2007). Single-unit firing in rat perirhinal cortex caused by fear conditioning to arbitrary and ecological stimuli. Journal of Neuroscience, 27, 12277-12291.
GAUB, S., GROSZER, M., FISHER, S.E. & EHRET, G. (2010). The structure of innate vocalizations in Foxp2-deficient mouse pups. Genes Brain and Behavior., 9, 390-401.
GROSZER, M., KEAYS, D.A., DEACON, R.M., DE BONO, J.P., PRASAD-MULCARE, S., GAUB, S., BAUM, M.G., FRENCH, C.A., NICOD, J., COVENTRY, J.A., ENARD, W., FRAY, M., BROWN, S.D., NOLAN, P.M., PÄÄBO, S., CHANNON, K.M., COSTA, R.M., EILERS, J., EHRET, G., RAWLINS, J.N. & FISHER, S.E. (2008). Impaired synaptic plasticity and motor learning in mice with a point mutation implicated in human speech deficits. Current Biology, 18, 354-362.
HAMMERSCHMIDT, K., RADYUSHKIN, K., EHRENREICH, H. & FISCHER, J. (2009). Female mice respond to male ultrasonic ‘songs’ with approach behaviour. Biology Letters, 5, 589-592.
HAMMERSCHMIDT, K., REISINGER, E., WESTEKEMPER, K., EHRENREICH, L., STRENZKE, N. & FISCHER, J. (2012). Mice do not require auditory input for the normal development of their ultrasonic vocalizations. BMC Neuroscience, 13, 40.
HARMON, K.M., CROMWELL, H.C., BURGDORF, J., MOSKAL, J.R., BRUDZYNSKI, S.M., KROES, R.A. & PANKSEPP, J. (2008). Rats selectively bred for low levels of 50 kHz ultrasonic vocalizations exhibit alterations in early social motivation. Developmental Psychobiology, 50, 322-331.
HIMMLER, B.T., KISKO, T.M., EUSTON, D.R., KOLB, B. & PELLIS, S.M. (2014). Are 50-kHz calls used as play signals in the playful interactions of rats? I. Evidence from the timing and context of their use. Behavioural Processes, 106, 60-66.
HOFER, M.A. (1996). Multiple regulators of ultrasonic vocalization in the infant rat. Psychoneuroendocrinology, 21, 203-217.
HOFER, M.A. & SHAIR, H. (1978). Ultrasonic vocalization during social interaction and isolation in 2-weeek-old rats. Developmental Psychobiology, 11, 495-504.
HOLY, T.E. & GUO, Z. (2005). Ultrasonic songs of male mice. PLoS Biol, 3, e386.
INSEL, T.R., HILL, J.L. & MAYOR, R.B. (1986). Rat pup ultrasonic isolation calls: possible mediation by the benzodiazepine receptor complex. Pharmacology Biochemistry and Behavior, 24, 1263-7.
JAMAIN, S., RADYUSHKIN, K., HAMMERSCHMIDT, K., GRANON, S., BORETIUS, S., VAROQUEAUX, F., RAMANANTSOA, N., GALLEGO, J., RONNENBERG, A., WINTER, D., FRAHM, J., FISCHER, J., BOURGERON, T., EHRENREICH, H. & BROSE, N. (2008). Reduced social interaction and ultrasonic communication in a mouse model of monogenic heritable autism. Proceedings of the National Academy of Sciences of the United States of America, 105, 1710-1715. JIANG, Y.H., PAN, Y., ZHU, L., LANDA, L., YOO, J., SPENCER, C., LORENZO, I., BRILLIANT, M., NOEBELS, J. & BEAUDET, A.L. (2010). Altered ultrasonic vocalization and impaired learning and memory in Angelman syndrome mouse model with a large maternal deletion from Ube3a to Gabrb3. PLoS One, 5, e12278.
KALTWASSER, M.T. (1990). Acoustic signaling in the black rat (Rattus rattus). Journal of Comparative Psychology, 104, 227-232.
KEHNE, J.H., COVERDALE, S., MCCLOSKEY, T.C., HOFFMAN, D.C. & CASSELLA, J.V. (2000). Effects of the CRF(1) receptor antagonist, CP 154,526, in the separation-induced vocalization anxiolytic test in rat pups. Neuropharmacology, 39, 1357-1367.
KESSLER, M.S., BOSCH, O.J., BUNCK, M., LANDGRAF, R. & NEUMANN, I.D. (2011). Maternal care differs in mice bred for high vs. low trait anxiety: impact of brain vasopressin and cross-fostering. Social Neuroscience, 6, 156-168.
KIM, E.J., KIM, E.S., COVEY, E. & KIM, J.J. (2010). Social Transmission of Fear in Rats: The Role of 22-kHz Ultrasonic Distress Vocalization. PLoS One, 5, e15077.
KISKO, T.M., HIMMLER, B.T., HIMMLER, S.M., EUSTON, D.R. & PELLIS, S.M. (2015). Are 50-kHz calls used as play signals in the playful interactions of rats? II. Evidence from the effects of devocalization. Behavioural Processes, 111, 25-33.
KNUTSON, B., BURGDORF, J. & PANKSEPP, J. (1998). Anticipation of play elicits high-frequency ultrasonic vocalizations in young rats. Journal of Comparative Psychology, 112, 65-73.
KNUTSON, B., BURGDORF, J. & PANKSEPP, J. (1999). High-frequency ultrasonic vocalizations index conditioned pharmacological reward in rats. Physiology and Behavior, 66, 639-643.
KRÖMER, S.A., KESSLER, M.S., MILFAY, D., BIRG, I.N., BUNCK, M., CZIBERE, L., PANHUYSEN, M., PÜTZ, B., DEUSSING, J.M., HOLSBOER, F., LANDGRAF, R. & TURCK, C.W. (2005). Identification of glyoxalase-I as a protein marker in a mouse model of extremes in trait anxiety. The Journal of Neuroscience, 25, 4375-4384.
KURZ, A., WÖHR, M., WALTER, M., BONIN, M., AUBURGER, G., GISPERT, S. & SCHWARTING, R.K.W. (2010). Alpha-synuclein deficiency affects brain Foxp1 expression and ultrasonic vocalization. Neuroscience, 166, 785-795.
LAVIOLA, G., ADRIANI, W., GAUDINO, C., MARINO, R. & KELLER, F. (2006). Paradoxical effects of prenatal acetylcholinesterase blockade on neuro-behavioral development and drug-induced stereotypies in reeler mutant mice. Psychopharmacology, 187, 331-344.
LUKAS, M. & WÖHR, M. (2015). Endogenous vasopressin, innate anxiety, and the emission of pro-social 50-kHz ultrasonic vocalizations during social play behavior in juvenile rats. Psychoneuroendocrinology, 56, 35-44.
MÄLLO, T., MATROV, D., HERM, L., KOIV, K., ELLER, M., RINKEN, A. & HARRO, J. (2007). Tickling-induced 50-kHz ultrasonic vocalization is individually stable and predicts behaviour in tests of anxiety and depression in rats. Behavioural Brain Research, 184, 57-71.
MAGGIO, J.C., MAGGIO, J.H. & WHITNEY, G. (1983). Experience-based vocalization of male mice to female chemosignals. Physiology & Behavior, 31, 269-272.
MAHRT, E.J., PERKEL, D.J., TONG, L., RUBEL, E.W. & PORTFORS, C.V. (2013). Engineered deafness reveals that mouse courtship vocalizations do not require auditory experience. The Journal of Neuroscience, 33, 5573-5583.
MAIER, E.Y., MA, S.T., AHRENS, A., SCHALLERT, T.J. & DUVAUCHELLE, C.L. (2010). Assessment of ultrasonic vocalizations during drug self-administration in rats. Journal of Visualized Experiments, 41, 2041.
MALKESMAN, O., SCATTONI, M.L., PAREDES, D., TRAGON, T., PEARSON, B., SHALTIEL, G., CHEN, G., CRAWLEY, J.N. & MANJI, H.K. (2010). The female urine sniffing test: a novel approach for assessing reward-seeking behavior in rodents. Biological Psychiatry, 67, 864-871.
MCGINNIS, M.Y. & VAKULENKO, M. (2003). Characterization of 50-kHz ultrasonic vocalizations in male and female rats. Physiology and Behavior, 80, 81-8.
MOLEWIJK, H.E., VAN DER POEL, A.M., MOS, J., VAN DER HEYDEN, J.A.M. & OLIVIER, B. (1995). Conditioned ultrasonic distress vocalizations in adult male rats as a behavioral paradigm for screening anti-panic drugs. Psychopharmacology, 117, 32-40.
MOSIENKO, M., BEIS, D., ALENINA, N. & WÖHR, M. (in press). Reduced isolation-induced pup ultrasonic communication in mouse pups lacking brain serotonin. Molecular Autism.
NAKATANI, J., TAMADA, K., HATANAKA, F., ISE, S., OHTA, H., INOUE, K., TOMONAGA, S., WATANABE, Y., CHUNG, Y.J., BANERJEE, R., IWAMOTO, K., KATO, T., OKAZAWA, M., YAMAUCHI, K., TANDA, K., TAKAO, K., MIYAKAWA, T., BRADLEY, A. & TAKUMI, T. (2009). Abnormal behavior in a chromosome-engineered mouse model for human 15q11-13 duplication seen in autism. Cell, 137, 1235-1246.
NASTITI, K., BENTON, D. & BRAIN, P.F. (1991). The effects of compounds acting at the benzodiazepine receptor complex on the ultrasonic calling of mouse pups. Behavioural Pharmacology, 2, 121-128.
NATUSCH, C. & SCHWARTING, R.K.W. (2010). Using bedding in a test environment critically affects 50-kHz ultrasonic vocalizations in laboratory rats. Pharmacology Biochemistry and Behavior, 96, 251-259.
NYBY, J., BIGELOW, J., KERCHNER, M. & BARBEHENN, F. (1983). Male mouse (Mus musculus) ultrasonic vocalizations to female urine: why is heterosexual experience necessary? Behavioral and Neural Biology, 38, 32-46.
NYBY, J., DIZINNO, G.A. & WHITNEY, G. (1976). Social status and ultrasonic vocalizations of male mice. Behavioral Biology, 18, 285-289.
NYBY, J., WYSOCKI, C.J., WHITNEY, G. & DIZINNO, G. (1977). Pheromonal regulation of male mouse ultrasonic courtship (Mus musculus). Animal Behaviour, 25, 333-341.
OLIVIER, B., MOLEWIJK, E., VAN OORSCHOT, R., VAN DER HEYDEN, J., RONKEN, E. & MOS, J. (1998a). Rat pup ultrasonic vocalization: effects of benzodiazepine receptor ligands. European Journal of Pharmacology, 358, 117-28.
OLIVIER, B., MOLEWIJK, H.E., VAN DER HEYDEN, J.A., VAN OORSCHOT, R., RONKEN, E., MOS, J. & MICZEK, K.A. (1998b). Ultrasonic vocalizations in rat pups: effects of serotonergic ligands. Neuroscience and Biobehavioral Reviews, 23, 215-27.
PANKSEPP, J. & BURGDORF, J. (2000). 50-kHz chirping (laughter?) in response to conditioned and unconditioned tickle-induced reward in rats: effects of social housing and genetic variables. Behavioural Brain Research, 115, 25-38.
PANKSEPP, J. & BURGDORF, J. (2003). “Laughing” rats and the evolutionary antecedents of human joy? Physiology and Behavior, 79, 533-547.
PANKSEPP, J., BURGDORF, J., TURNER, C. & GORDON, N. (2002). Modeling ADHD-type arousal with unilateral frontal cortex damage in rats and beneficial effects of play therapy. Brain and Cognition, 52, 97-105.
PANKSEPP, J.B., JOCHMAN, K.A., KIM, J.U., KOY, J.J., WILSON, E.D., CHEN, Q., WiILSON, C.R. & LAHVIS, G.P. (2007). Affiliative behavior, ultrasonic communication and social reward are influenced by genetic variation in adolescent mice. PLoS One, 2, e351.
PARSANA, A.J., LI, N. & BROWN, T.H. (2012a). Positive and negative ultrasonic social signals elicit opposing firing patterns in rat amygdala. Behavioural Brain Research, 226, 77-86.
PARSANA, A.J., MORAN, E.E. & BROWN, T.H. (2012b). Rats learn to freeze to 22-kHz ultrasonic vocalizations through autoconditioning. Behavioural Brain Research, 232, 395-399.
PEREIRA, M., ANDREATINI, R., SCHWARTIN, R.K.W. & BRENES, J.C. (2014) Amphetamine-induced appetitive 50-kHz calls in rats: a marker of affect in mania? . Psychopharmacology, 231, 2567-2577.
POMERANTZ, S.M., NUNEZ, A.A. & BEAN, N.J. (1983). Female behavior is affected by male ultrasonic vocalizations in house mice. Physiology and Behavior, 31, 91-96.
PORTFORS, C.V. & PERKEL, D.J. (2014). The role of ultrasonic vocalizations in mouse communication. Current Opinion in Neurobiology, 28, 115-120.
RADYUSHKIN, K., HAMMERSCHMIDT, K., BORETIUS, S., VAROQUEAUX, F., EL-KORDI, A., RONNENBERG, A., WINTER, D., FRAHM, J., FISCHER, J., BROSE, N. & EHRENREICH, H. (2009). Neuroligin-3-deficient mice: model of a monogenic heritable form of autism with an olfactory deficit. Genes Brain and Behavior, 8, 416-425.
ROULLET, F.I., WÖHR, M. & CRAWLEY, J.N. (2011). Female urine-induced male mice ultrasonic vocalizations, but not scent-marking, is modulated by social experience. Behavioural Brain Research, 216, 19-28.
RYGULA, R., GOLEBIOWSKA, J., KREGIEL, J., KUBIK, J. & POPIK, P. (2015). Effects of optimism on motivation in rats. Frontiers in Behavioural Neuroscience, 9, 32.
RYGULA, R., PLUTA, H. & POPIK, P. (2012). Laughing rats are optimistic. PLoS One, 7, e51959.
SADANANDA, M., WÖHR, M. & SCHWARTING, R.K.W. (2008). Playback of 22-kHz and 50-kHz ultrasonic vocalizations induces differential c-fos expression in rat brain. Neuroscience Letters, 435, 17-23.
SALES, G.D. & PYE, D. (1974). Ultrasonic communication by animals. New York: Wiley.
SCATTONI, M.L., CRAWLEY, J. & RICCERI, L. (2009). Ultrasonic vocalizations: a tool for behavioural phenotyping of mouse models of neurodevelopmental disorders. Neuroscience and Biobehavioral Reviews, 33, 508-515.
SCATTONI, M.L., GANDHY, S.U., RICCERI, L. & CRAWLEY, J.N. (2008). Unusual repertoire of vocalizations in the BTBR T+tf/J mouse model of autism. PLoS One, 3, e3067.
SCATTONI, M.L., MARTIRE, A., CARTOCCI, G., FERRANTE, A. & RICCERI, L. (2013). Reduced social interaction, behavioural flexibility and BDNF signalling in the BTBR T+ tf/J strain, a mouse model of autism. Behavioural Brain Research, 251, 35-40.
SCHMEISSER, M.J., EY, E., WEGENER, S., BOCKMANN, J., STEMPEL, A.V., KUEBLER, A., JANSSEN, A.L., UDVARDI, P.T., SHIBAN, E., SPILKER, C., BALSCHUN, D., SKRYABIN, B.V., DIECK, S.T., SMALLA, K.H., MONTAG, D., LEBLOND, C.S., FAURE, P., TORQUET, N., LE SOURD, A.M., TORO, R., GRABRUCKER, A.M., SHOICHERT, S.A., SCHMITZ, D., KREUTZ, M.R., BOURGERON, T., GUNDELFINGER, E.D. & BOECKERS, T.M. (2012). Autistic-like behaviours and hyperactivity in mice lacking ProSAP1/Shank2. Nature, 486, 256-260.
SCATTONI, M.L., RICCERI, L. & CRAWLEY, J.N. (2011). Unusual repertoire of vocalizations in adult BTBR T+tf/J mice during three types of social encounters. Genes Brain and Behavior.
SCHWARTING, R.K.W., JEGAN, N. & WÖHR, M. (2007). Situational factors, conditions and individual variables which can determine ultrasonic vocalizations in male adult Wistar rats. Behavioural Brain Research, 182, 208-222
SEFFER, D., RIPPBERGER, H., SCHWARTING, R.K.W. & WÖHR, M. (in press). Pro-social 50-kHz ultrasonic communication in rats: Post-weaning but not post-adolescent social isolation leads to social impairments – phenotypic rescue by re-socialization. Frontiers in Behavioral Neuroscience.
SEWELL, G.D. (1967). Ultrasound in adult rodents. Nature, 215, 512.
SEWELL, G.D. (1970). Ultrasonic communication in rodents. Nature, 227, 410.
SHU, W., CHO, J.Y., JIANG, Y., ZHANG, M., WEISZ, D., ELDER, G.A., SCHMEIDLER, J., DE GASPERI, R., SOSA, M.A., RABIDOU, D., SANTUCCI, A.C., PERL, D., MORRISEY, E.& BUXBAUM, J.D. (2005). Altered ultrasonic vocalization in mice with a disruption in the Foxp2 gene. Proceedings of the National Academy of Sciences of the United States of America, 102, 9643-9648.
SILVERMAN, J.L., YANG, M., LORD, C. & CRAWLEY, J.N. (2010). Behavioural phenotyping assays for mouse models of autism. Nature Reviews Neuroscience, 11, 490-502.
SIPOS, M.L., KERCHNER, M. & NYBY, J.G. (1992). An ephemeral sex pheromone in the urine of female house mice (Mus domesticus). Behavioral and Neural Biology, 58, 138-143.
SIPOS, M.L., WYSOCKI, C.J., NYBY, J.G., WYSOCKI, L. & NEMURA, T.A. (1995). An ephemeral pheromone of female house mice: perception via the main and accessory olfactory systems. Physiology and Behavior, 58, 529-534.
SMITH, J.C. (1976). Responses to adult mice to models infant calls. Journal of Comparative and Physiological Psychology, 90, 1105-1115.
SMOTHERMAN, W.P., BELL, R.W., STARZEC, J., ELIAS, J. & ZACHMAN, T.A. (1974). Maternal responses to infant vocalizations and olfactory cues in rats and mice. Behavioral Biology, 12, 55-66.
SCHWARTING, R.K.W., JEGAN, N. & WÖHR, M. (2007). Situational factors, conditions and individual variables which can determine ultrasonic vocalizations in male adult Wistar rats. Behavioural Brain Research, 182, 208-222.
SIVIY, S.M. &, PANKSEPP, J. (1987). Sensory modulation of juvenile play in rats. Developmental Psychobiology, 20, 39-55.
TAKAHASHI, A., YAP, J.J., BOHAGER, D.Z., Faccidomo, S., CLAYTON, T., COOK, J.M. & MICZEK, K.A. (2009). Glutamatergic and GABAergic modulations of ultrasonic vocalizations during maternal separation distress in mouse pups. Psychopharmacology, 204, 61-71.
VAN DER POEL, A.M., NOACH, E.J.K. & MICZEK, K.A. (1989). Temporal patterning of ultrasonic distress calls in the adult rat: effects of morphine and benzodiazepines. Psychopharmacology, 97, 147-148.
VANDERSCHUREN, L.J.L.J., NIESINK, R.J. & VAN REE, J.M. (1997). The neurobiology of social play behavior in rats. Neuroscience and Biobehavioral Reviews, 21, 309-26.
VIVIAN, J.A., BARROS, H.M., MANITIU, A. & MICZEK, K.A. (1997). Ultrasonic vocalizations in rat pups: modulation at the gamma-aminobutyric acidA receptor complex and the neurosteroid recognition site. The Journal of Pharmacology and Experimental Therapeutics, 282, 318-325.
VIVIAN, J.A. & MICZEK, K.A. (1993). Morphine attenuates ultrasonic vocalization during agonistic encounters in adult male rats. Psychopharmacology, 111, 367-375.
WHITE, N.R., CAGIANO, R., MOISES, A.U. & BARFIELD, R.J. (1990). Changes in mating vocalizations over the ejaculatory series in rats (Rattus norvegicus). Journal of Comparative Psychology, 104, 255-262.
WHITNEY, G., ALPERN, M., DIZINNO, G. & HOROWITZ, G. (1974). Female odors evoke ultrasounds from male mice. Animal Learning and Behavior, 2, 13-18.
WIGGER, A., LOERSCHER, P., WEISSENBACHER, P., HOLSBOER, F. & LANDGRAF, R. (2001). Cross-fostering and cross-breeding of HAB and LAB rats: a genetic rat model of anxiety. Behavior Genetics, 31, 371-382.
WILLADSEN, M., SEFFER, D., SCHWARTING, R.K.W. & WÖHR, M. (2014). Rodent ultrasonic communication: Male prosocial 50-kHz ultrasonic vocalizations elicit social approach behavior in female rats (Rattus norvegicus). Journal of Comparative Psychology, 128, 56-64.
WILLUHN, I., TOSE, A., WANAT, M.J., HART, A.S., HOLLON, N.G., PHILLIPS, P.E.M., SCHWARTING, R.K.W. & WÖHR, M. (2014). Phasic dopamine release in the nucleus accumbens in response to pro-social 50 kHz ultrasonic vocalizations in rats. The Journal of Neuroscience, 34, 10616-10623.
WINSLOW, J.T. & INSEL, T.R. (2002). The social deficits of the oxytocin knockout mouse. Neuropeptides, 36, 221-229.
WINTINK, A.J. & BRUDZYNSKI, S.M. (2001). The related roles of dopamine and glutamate in the initiation of 50-kHz ultrasonic calls in adult rats. Pharmacology Biochemistry and Behavior, 70, 317-323.
WRIGHT, J.M., GOURDON, J.C. & CLARKE, P.B. (2010). Identification of multiple call categories within the rich repertoire of adult rat 50-kHz ultrasonic vocalizations: effects of amphetamine and social context. Psychopharmacology, 211, 1-13.
WÖHR, M. (2014). Ultrasonic vocalizations in Shank mouse models for autism spectrum disorders: detailed spectrographic analyses and developmental profiles. Neuroscience and Biobehavioral Reviews, 43, 199-212.
WÖHR, M. (2015). Effect of social odor context on the emission of isolation-induced ultrasonic vocalizations in the BTBR T+tf/J mouse model for autism. Frontiers in Neuroscience, 9, 73.
WÖHR, M., BORTA, A. & SCHWARTING, R.K.W. (2005). Overt behavior and ultrasonic vocalization in a fear conditioning paradigm: A dose-response study in the rat. Neurobiology of Learning and Memory, 84, 228-240.
WÖHR, M., DAHLHOFF, M., WOLF, E., HOLSBOER, F., SCHWARTING, R.K.W. & WOTJAK, C.T. (2008a). Effects of genetic background, gender, and early environmental factors on isolation-induced ultrasonic calling in mouse pups: an embryo-transfer study. Behavior Genetics, 38, 579-595.
WÖHR, M., HOUX, B., SCHWARTING, R.K.W. & SPRUIJT, B. (2008b). Effects of experience and context on 50-kHz vocalizations in rats. Physiology and Behavior, 93, 766-776.
WÖHR, M., MOLES, A., SCHWARTING, R.K.W. & D’AMATO, F.R. (2011a) Lack of social exploratory activation in male mu-opioid receptor KO mice in response to playback of female ultrasonic vocalizations. Social Neuroscience, 6, 76-87.
WÖHR, M., KEHL, M., BORTA, A., SCHÄNZER, A., SCHWARTING, R.K.W. & HÖGLINGER, G.U. (2009). New insights into the relationship of neurogenesis and affect: tickling induces hippocampal cell proliferation in rats emitting appetitive 50-kHz ultrasonic vocalizations. Neuroscience, 163, 1024-1030.
WÖHR, M., ROULLET, F.I. & CRAWLEY, J.N. (2011b). Reduced scent marking and ultrasonic vocalizations in the BTBR T+tf/J mouse model of autism. Genes Brain and Behavior, 10, 35-43.
WÖHR, M. & SCHWARTING, R.K.W. (2007). Ultrasonic communication in rats: can playback of 50-kHz calls induce approach behavior? PLoS One, 2, e1365.
WÖHR, M. & SCHWARTING, R.K.W. (2008a). Maternal care, isolation-induced infant ultrasonic calling, and their relations to adult anxiety-related behavior in the rat. Behavioral Neuroscience, 122, 310-330.
WÖHR, M. & SCHWARTING, R.K.W. (2008b). Ultrasonic calling during fear conditioning in the rat: No evidence for an audience effect. Animal Behaviour, 76, 749-760.
WÖHR, M. & SCHWARTING, R.K.W. (2009). Ultrasonic communication in rats: effects of morphine and naloxone on vocal and behavioral responses to playback of 50-kHz vocalizations. Pharmacology Biochemistry and Behavior, 94, 285-295.
WÖHR, M. & SCHWARTING, R.K.W. (2010). Activation of limbic system structures by replay of ultrasonic vocalization in rats. In: Handbook of Mammalian Vocalization. An Integrative Neuroscience Approach. Brudzynski, S.M. (Ed.). Elsevier.
WÖHR, M. & SCHWARTING, R.K.W. (2013). Affective communication in rodents: ultrasonic vocalizations as a tool for research on emotion and motivation. Cell and Tissue Research, 354, 81-97.
YAMAMURO, T., SENZAKI, K., IWAMOTO, S., NAKAGAWA, Y., HAYASHI, T., HORI, M., SAKAMOTO, S., MURAKAMI, K., SHIGA, T. & URAYAMA, O. (2010). Neurogenesis in the dentate gyrus of the rat hippocampus enhanced by tickling stimulation with positive emotion. Neuroscience Research, 68, 285-289.
YEE, N., SCHWARTING, R.K.W., FUCHS, E. & WÖHR, M. (2012a). Juvenile stress potentiates aversive 22-kHz ultrasonic vocalizations and freezing during auditory fear conditioning in adult male rats. Stress, 15, 533-544.
YEE, N., SCHWARTING, R.K.W., FUCHS, E. & WÖHR, M. (2012b). Increased affective ultrasonic communication during fear learning in adult male rats exposed to maternal immune activation. Journal of Psychiatric Research, 46, 1199-11205.
ZIPPELIUS, H.M. & SCHLEIDT, W.M. (1956). Ultraschall-Laute bei jungen Mäusen. Naturwissenschaften, 43, 502.
Correspondence should be addressed to:
Dr. Markus Wöhr
Behavioural Neuroscience
Experimental and Physiological Psychology
Philipps-University of Marburg
Gutenbergstr. 18, 35032 Marburg, Germany
Fax: +6421-282 3610, Tel: +6421-28 23612
e-mail: markus.woehr@staff.uni-marburg.de
Websites:
Team Schwarting
AG Wöhr – Philipps-Universität Marburg – Fb. 04 – Psychologie
Examples of rat ultrasonic vocalizations (USV)
Norwegian Rat (Rattus norvegicus), Wistar albino strain, Males
The following recordings have been made with Avisoft-UltraSoundGate 116-200 at a sample rate of 214.285 kHz. In order to make the recordings audible, the sample rate of the original wav files has been set to 11.025 kHz (providing a time expansion factor of 19.44).
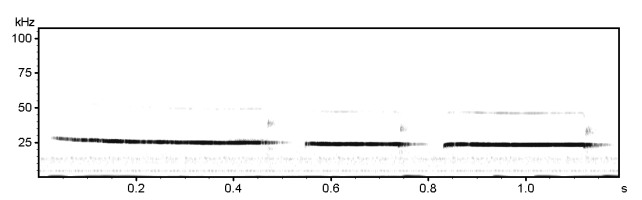
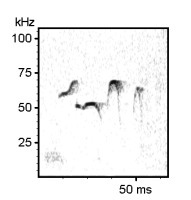
22 kHz rat calls (‘distress’ calls). Listen to the time-expanded wav file.

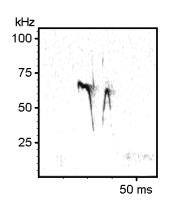
40 kHz rat calls (note the different time scales of the two spectrograms). Listen to the time-expanded wav file.
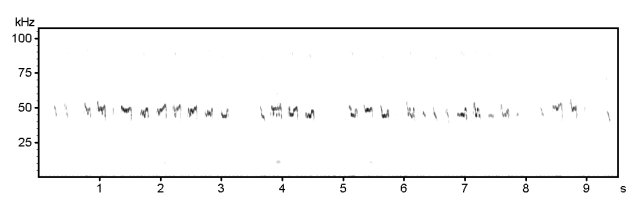
Various 50 kHz rat calls :
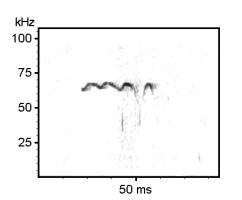
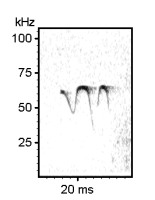


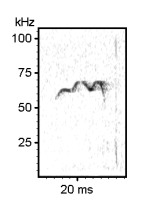
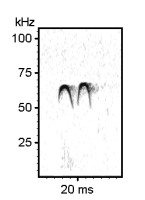
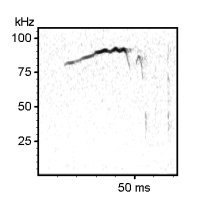
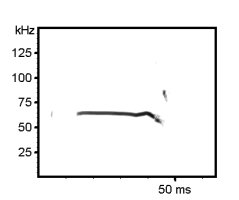
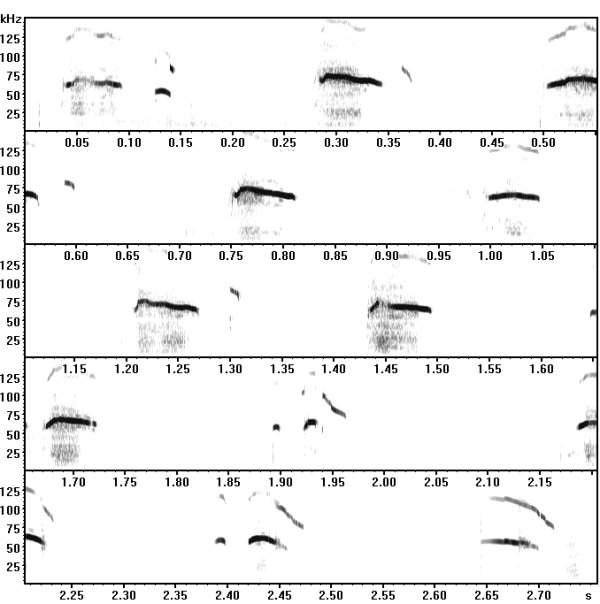
Examples of mouse ultrasonic vocalizations (USV)
Laboratory mouse (Mus musculus), C57BL/6JOlaHSd strain, infant male, 7 days old, separation-induced vocalizations
The following recordings have been made with Avisoft-UltraSoundGate 116-200 at a sample rate of 300 kHz. In order to make the recordings audible, the sample rate of the original wav files has been set to 22.05 kHz (providing a time expansion factor of 13.6).



All sound recordings by Markus Wöhr (Philipps-University of Marburg, Germany).
An overview on the Avisoft Bioacoustics products suited for recording and analyzing mice and rat USV is available at the USV products overview page.